海洋與生態(進階):養分、缺氧與藻華開關的隱形動力學
從 Redfield 比值、鐵限制與 Sverdrup 臨界深度,到微生物迴路與含氧最小帶,潛入海洋生產力背後的化學計量與物理耦合
同樣有陽光、同樣有水,為什麼有些藍色海域永遠長不出魚?
讀過入門篇,你已經知道海洋靠浮游植物固碳、能量沿食物網一層層遞減、湧升流養出豐饒漁場。但這裡有個讓海洋學家困惑了數十年的謎題:南冰洋(Southern Ocean)、赤道東太平洋、亞北極太平洋這些海域,表層的氮、磷一年到頭都用不完,陽光也充足,照理說應該綠油油一片,實際上浮游植物卻稀稀落落、葉綠素濃度偏低。它們被稱為「高營養鹽、低葉綠素」海區(high-nutrient, low-chlorophyll, HNLC)。
養分有、光也有,浮游植物卻不長——這代表入門篇「生產力受光與養分共同限制」的說法,還缺了一塊拼圖。這一篇進階文章,我們不再重講分層與食物網,而是直接潛入三個更硬核的問題:浮游植物到底「缺什麼」(養分限制的化學計量學)、春天大爆發為什麼會在某個深度突然啟動(Sverdrup 臨界深度模型)、以及海洋裡其實藏著一條入門篇沒提的隱形迴路(微生物迴路與溶解有機碳)。最後,我們會看暖化如何透過「缺氧」這條路,悄悄改寫整片海洋的代謝地圖。
化學計量學:浮游植物不是想長就能長
入門篇說生產力受氮、磷限制。但「限制」具體是什麼意思?這要從一個漂亮的觀察說起。1930 年代,海洋學家 Alfred Redfield 發現:全球海洋裡浮游植物體內,以及溶解在深層海水中的碳、氮、磷,比例驚人地穩定,接近
$$\mathrm{C} : \mathrm{N} : \mathrm{P} = 106 : 16 : 1$$
這就是著名的 Redfield 比值(Redfield ratio)。它的深意在於:浮游植物「組裝」自己的身體需要固定配方的原料。把光合作用與呼吸合在一起寫,海洋生物的生成與再礦化可表示為:
$$106\,\mathrm{CO_2} + 16\,\mathrm{HNO_3} + \mathrm{H_3PO_4} + 122\,\mathrm{H_2O} \rightleftharpoons (\mathrm{C_{106}H_{263}O_{110}N_{16}P_1}) + 138\,\mathrm{O_2}$$
向右是合成有機物(生產),向左是分解再礦化(呼吸)。注意右邊釋放 $138$ 莫耳的 $\mathrm{O_2}$——這個數字之後談缺氧時會回來找我們。
有了固定配方,「誰是限制因子」就變成一個化學計量問題。這正是 Liebig 最小因子律(Liebig's law of the minimum) 在海洋裡的版本:生長速率由「相對於需求最稀缺」的那個養分決定,而不是絕對濃度最低的那個。把它寫成判斷式:比較海水中可用氮磷的實際比值與 Redfield 比值。若 $\mathrm{N}:\mathrm{P} < 16$,氮相對不足,系統由氮限制;若 $\mathrm{N}:\mathrm{P} > 16$,則磷限制。大洋多半是氮限制(這也是固氮藍綠菌如 Trichodesmium 重要的原因),而許多湖泊與部分近岸是磷限制。
那 HNLC 海區到底缺什麼?
回到開頭的謎題。HNLC 海區氮磷明明過剩,按 Liebig 律應該不缺。1980 年代 John Martin 提出大膽假設:它們缺的是鐵(Fe)。鐵是光合作用電子傳遞鏈、硝酸還原酶的必需微量元素,需求量極小,但開放大洋遠離陸源塵土輸入,鐵濃度可低到 picomolar 等級,反而成了真正的「最小因子」。Martin 留下一句名言:「給我半船鐵,我就還你一個冰期。」
後續的中尺度加鐵實驗(如 IronEx、SOFeX)證實了這點:在 HNLC 海域人為撒入可溶性鐵後,浮游植物在數天內爆發、葉綠素飆升、表層 $\mathrm{CO_2}$ 下降。這把入門篇的「養分限制」推進到一個更精細的層次——限制因子是相對的、是多元素的,而且常常輪到不起眼的微量金屬出場。對台灣周邊而言,東亞沙塵(Asian dust)的沉降正是西北太平洋一個重要的鐵與營養鹽來源,沙塵事件之後常可在衛星影像上看到葉綠素的短暫上升。
Sverdrup 臨界深度:春天大爆發的開關藏在哪裡
入門篇談湧升流如何把養分送上來,但溫帶與高緯海域還有另一個壯觀現象:春季藻華(spring bloom)——每年春天浮游植物在幾週內突然爆量。為什麼是春天?為什麼會「突然」啟動?答案是 1953 年 Harald Sverdrup 提出的臨界深度理論(critical depth hypothesis),它是海洋生物物理耦合最經典的模型之一。
關鍵在於浮游植物會被海水的湍流混合,在混合層(mixed layer) 裡上下翻攪。一個細胞時而被帶到光亮的表層(光合作用 > 呼吸,淨賺),時而被帶到昏暗的深處(光合作用 < 呼吸,淨虧)。整群浮游植物能否淨增長,取決於它們在混合層裡「平均下來」是賺還是賠。
定義兩個深度:
- 補償深度(compensation depth) $z_c$:在此深度,單一細胞的光合作用速率恰等於呼吸速率,光強約為表面的 $1\%$。
- 臨界深度(critical depth) $z_{cr}$:在此深度之上,整個水柱積分的光合作用總量恰等於整個水柱積分的呼吸總量。
藻華啟動的條件是:
$$z_{\text{mix}} < z_{cr}$$
也就是混合層深度 $z_{\text{mix}}$ 要淺於臨界深度。冬天風強、表層冷卻劇烈,混合層可深達數百公尺,浮游植物大半時間待在黑暗中,群體淨虧損,長不起來。到了春天,日照增強、表層回暖,水體分層(stratification) 形成,混合層快速變淺;一旦 $z_{\text{mix}}$ 跌破 $z_{cr}$,浮游植物平均待在亮處的時間夠長,族群淨增長轉正——藻華「啪」地一聲啟動。
動手算一下:估一個臨界深度
臨界深度 $z_{cr}$ 滿足「水柱積分光合作用 = 水柱積分呼吸」。在常見的簡化假設下(光強隨深度指數衰減 $I(z)=I_0 e^{-kz}$、呼吸速率沿深度近似定值),可推得 Sverdrup 的近似式。當 $k\,z_{cr}$ 夠大時,
$$z_{cr} \approx \frac{I_0}{k \cdot I_c}$$
其中 $I_0$ 是海面有效光強、$I_c$ 是補償光強(光合=呼吸時的光強)、$k$ 是消光係數。我們代入一組合理的春季數值:設 $I_0 / I_c = 40$(表面光遠強於補償光),消光係數 $k = 0.1\ \mathrm{m^{-1}}$(清澈大洋)。
$$z_{cr} \approx \frac{40}{0.1\ \mathrm{m^{-1}}} = 400\ \mathrm{m}$$
這告訴我們:只要春天的混合層淺於約 $400$ 公尺,藻華就會啟動。冬季混合層若深達 $500$ 公尺,則被壓在臨界深度之下,藻華按兵不動。把數字換一下你就明白為什麼近岸混濁水($k$ 大)的臨界深度淺得多——例如 $k=0.4\ \mathrm{m^{-1}}$ 時 $z_{cr}\approx 100$ 公尺,藻華更難啟動,需要更強的分層配合。
值得補充的是,現代研究(如 Behrenfeld 的「稀釋-再耦合假說」)指出,Sverdrup 模型忽略了浮游動物的攝食壓力,真實藻華的啟動有時在冬季混合最深、捕食者與獵物被「稀釋」脫鉤時就已悄悄醞釀。但臨界深度作為一個把光、混合、呼吸三者用一條不等式串起來的思想框架,至今仍是理解季節性生產力的基石。
微生物迴路:入門食物網漏掉的半個海洋
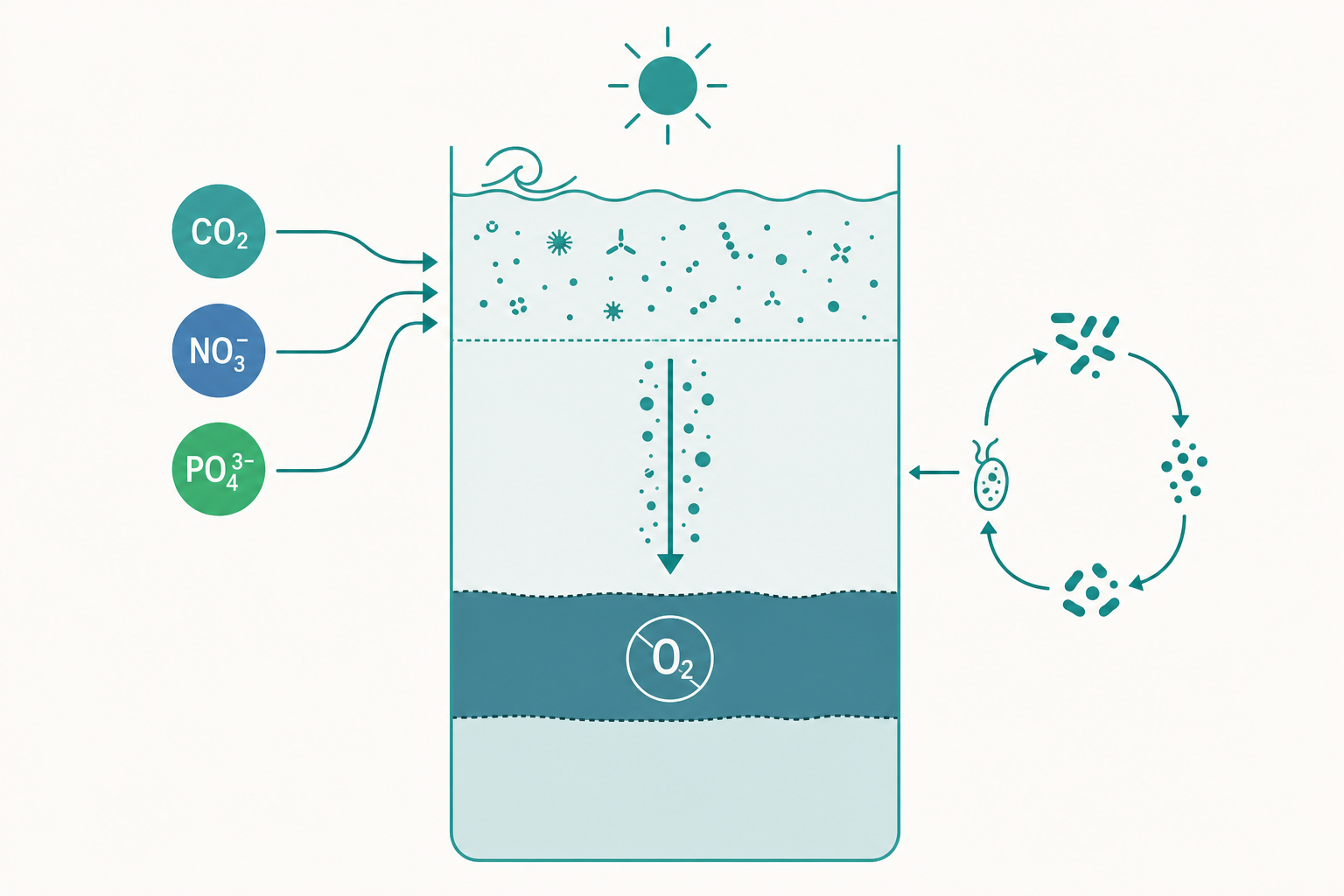
入門篇畫的食物網是「浮游植物→浮游動物→小魚→大魚」。但這條鏈條,其實漏掉了海洋裡數量最龐大、代謝最活躍的一群玩家——細菌,以及它們賴以維生的溶解有機碳(dissolved organic carbon, DOC)。
事情是這樣的:浮游植物固定的碳,並不會全部變成「顆粒」被浮游動物吃掉。其中有相當比例(常達 $30\%$ 以上)以溶解態的形式滲漏到海水中——可能來自細胞滲漏、病毒裂解破胞、浮游動物進食時的「邋遢取食(sloppy feeding)」、或排泄。這些溶解有機物太小、太稀,浮游動物根本濾不到、吃不著,傳統食物鏈視角下它們等於「漏掉的能量」。
1983 年 Azam 等人提出微生物迴路(microbial loop) 補上了這一塊:
$$\text{浮游植物} \rightarrow \text{DOC} \rightarrow \text{異營細菌} \rightarrow \text{原生動物(鞭毛蟲/纖毛蟲)} \rightarrow \text{浮游動物}$$
異營細菌(heterotrophic bacteria)能吸收溶解有機碳、把它重新組裝回顆粒態的「菌體生物量」,再被微小的原生動物捕食,原生動物又被浮游動物吃掉。於是原本要漏掉的溶解碳,透過微生物這個「迴路」被回收進主食物網。海洋因此不是一條乾淨的鏈,而是一張鏈與迴路交織的網。
這個迴路有兩個深遠後果。其一,它解釋了為什麼貧養大洋(「海洋沙漠」)雖然魚不多,微生物活性卻不低——能量大量在看不見的微生物層級空轉。其二,它連結到微生物碳泵(microbial carbon pump) 的概念:細菌反覆加工後,會留下一批極難被分解的惰性溶解有機碳(refractory DOC, RDOC),這些碳可以在深海漂流數千年不被礦化。海洋 DOC 庫的總碳量約 $662$ 億噸(Gt C),與整個大氣的碳量同一量級——這是一個入門篇完全沒提、卻舉足輕重的巨大碳庫。
缺氧的海洋:當呼吸追上供氧
最後我們把化學計量學與生物泵的線索接起來,看一個正在惡化的全球現象:海洋去氧化(ocean deoxygenation)。
還記得 Redfield 方程式右邊那 $138$ 莫耳 $\mathrm{O_2}$ 嗎?它揭示一個冷酷的對稱:生產有機物時放出多少氧,分解同樣多的有機物就要耗掉等量的氧。透光帶有光合作用持續造氧、又與大氣交換,所以氧氣充足;但在中層水,沉降下來的有機物被微生物再礦化,呼吸不斷抽走溶氧,又沒有光合作用補回——於是形成了含氧最小帶(oxygen minimum zone, OMZ),通常位於約 $200$ 至 $1000$ 公尺的深度。
OMZ 的存在與否,是「供氧」與「耗氧」的賽跑:供氧靠物理輸送(溶解度、環流通風),耗氧靠生物泵的輸出生產力。我們可以用一個簡單的質量平衡來把握它:
$$\frac{d[\mathrm{O_2}]}{dt} = \underbrace{\text{物理供氧}}_{\text{通風 + 擴散}} - \underbrace{r_{\mathrm{O_2}} \cdot R_{\text{remin}}}_{\text{再礦化耗氧}}$$
其中 $R_{\text{remin}}$ 是有機碳再礦化速率,$r_{\mathrm{O_2}}$ 是再礦化每單位碳的耗氧比(由 Redfield 化學計量決定,約 $138/106\approx 1.3$)。當供氧跟不上耗氧,溶氧持續下探,[O₂] 趨近於零,這個區域就對需氧生物變得不適合居住。
暖化會從兩端同時惡化這個平衡:一方面,氧氣在較暖的海水中溶解度下降(暖水裝不下那麼多氧);另一方面,暖化加劇表層分層,削弱垂直通風,深層更難補氧。兩者疊加,使全球 OMZ 在過去半世紀持續擴張。對漁業的衝擊很直接——鮪魚、旗魚等高耗氧的掠食者會被壓縮到表層較薄的含氧水層,棲地被「擠扁」;而一些能耐低氧的物種則趁勢擴張,群聚結構隨之重組。把這條線索拉回台灣:西北太平洋與南海邊緣同樣面臨增溫與分層增強,理解 OMZ 的動力學,正是預測未來漁場垂直分布與物種更替的關鍵之一。
重點回顧
- Redfield 比值 $\mathrm{C}:\mathrm{N}:\mathrm{P}=106:16:1$ 是浮游植物的「組裝配方」;配合 Liebig 最小因子律,可用實際 $\mathrm{N}:\mathrm{P}$ 偏離 $16$ 來判斷氮限制或磷限制。
- HNLC 海區氮磷過剩卻不長藻,真正的限制因子是微量元素鐵(Fe);加鐵實驗證實撒鐵能觸發藻華,限制因子是相對而非絕對的。
- Sverdrup 臨界深度理論用 $z_{\text{mix}} < z_{cr}$ 一條不等式解釋春季藻華的啟動:春天分層使混合層變淺、跌破臨界深度,浮游植物群體淨增長轉正。
- 微生物迴路把溶解有機碳(DOC)經由細菌、原生動物回收進主食物網,並衍生出與大氣同量級的巨大海洋 DOC 碳庫與微生物碳泵。
- 含氧最小帶(OMZ) 是供氧與生物泵耗氧的賽跑結果;暖化從溶解度下降與分層增強兩端惡化平衡,使 OMZ 擴張、壓縮高耗氧魚類棲地。
深入探討(研究所視角)
從固定配方到可變化學計量:Redfield 比值不是常數
研究所層級必須鬆開 Redfield 比值「處處 $106:16:1$」的假設。實際上細胞的 $\mathrm{C}:\mathrm{N}:\mathrm{P}$ 會隨環境與族群可塑性變動,這由生態化學計量學(ecological stoichiometry) 處理。一個關鍵框架是「生長速率假說(growth rate hypothesis)」:快速生長的細胞需要大量富磷的核糖體 RNA 來支撐蛋白質合成,因此高生長速率對應較低的 $\mathrm{C}:\mathrm{P}$ 與 $\mathrm{N}:\mathrm{P}$。反之,養分匱乏、生長受限的群落會囤積碳、提高 $\mathrm{C}:\mathrm{P}$。
這個可變性對碳循環有放大效應。若貧養化海域的浮游植物以更高的 $\mathrm{C}:\mathrm{P}$ 固碳(每消耗一個磷帶走更多碳),則同樣的養分供給能輸出更多碳到深海。將可變化學計量耦合進地球系統模式(Earth System Model),是目前估計生物泵對氣候回饋的活躍前沿,也是 CMIP 模式間離散度的重要來源之一。
把限制因子推到極致:共限制與「鐵-光-氮」三角
Liebig 律的「單一最小因子」是個一階近似。現場常見的是共限制(co-limitation):兩種以上資源同時逼近不足,移除其一仍不足以解除限制。海洋學界區分至少三類——獨立的生化共限制(不同元素用於不同酶系統)、生化替代共限制(如鈷可部分替代鋅)、以及鐵-光共限制。後者特別關鍵:光合電子傳遞鏈本身富含鐵蛋白,光照越弱、細胞需要越多反應中心,鐵需求越高。因此在深混合層(光受限)的 HNLC 海域,鐵與光會交織成耦合限制,這也回頭與 Sverdrup 臨界深度模型對話——混合層深度同時透過「平均光照」與「鐵需求」兩條路徑影響生產力。把 HNLC、鐵限制、臨界深度三者統一在一個資源競爭框架下,是生物海洋學一個優雅的整合方向。
臨界深度模型的數學內核
Sverdrup 近似式 $z_{cr}\approx I_0/(k I_c)$ 背後其實是一條積分平衡。設淨生長率密度為光合作用減呼吸,水柱積分平衡寫成
$$\int_0^{z_{cr}} P_{\max}\big(1-e^{-I(z)/I_k}\big)\,dz = \int_0^{z_{cr}} R\,dz$$
左邊用飽和型光合作用-光照曲線(P-I curve),右邊為定值呼吸 $R$。在弱光近似 $I(z)/I_k \ll 1$ 下線性化,並代入 $I(z)=I_0 e^{-kz}$ 積分,即可解出 $z_{cr}$ 與 $I_0$、$k$、$R$ 的關係,得到前述反比形式。這個推導的價值在於:它讓「藻華開關」不再是定性敘述,而是一個可被觀測量(衛星海表光、混合層深度 MLD、消光係數)參數化、可驗證的判據。當代用 Argo 浮標的 MLD 與遙測葉綠素去檢驗臨界深度與稀釋-再耦合兩派假說,正是把這條積分式拉到全球尺度做檢定。
連結與展望
把四條線索收束起來:浮游植物的化學計量決定了它「需要什麼、固定多少碳」;鐵與共限制決定了「哪裡長得起來」;臨界深度決定了「何時啟動」;而再礦化的化學計量又決定了深層「耗掉多少氧」。它們在一個耦合系統裡彼此牽動——暖化加強分層,既改變臨界深度與藻華時序,又削弱通風、擴張 OMZ,還可能透過貧養化推升 $\mathrm{C}:\mathrm{P}$ 而改變碳輸出效率。對研究者而言,下一步是把生態化學計量、微生物迴路與物理混合,整合進能預測區域差異的地球系統模式,並用 Argo、BGC-Argo、衛星遙測與基因體資料交叉驗證。生物海洋學、生物地球化學與物理海洋學在此會合——而台灣鄰接黑潮、南海與西北太平洋的特殊地理位置,正是觀測這些耦合過程如何在區域尺度上展開的天然實驗室。